Митохондрии - органеллы энергообеспечения метаболических процесов в клетке. Размеры их варьируют от 0,5 до 5-7 мкм, количество в клетке составляет от 50 до 1000 и более. В гиалоплазме митохондрии распределены обычно диффузно, однако в специализированных клетках сосредоточены в тех участках, где имеется наибольшая потребность в энергии. Например, в мышечных клетках и симпластах большие количества митохондрий сосредоточены вдоль рабочих элементов - сократительных фибрилл. В клетках, функции которых сопряжены с особо высокими энергозатратами, митохондрии образуют множественные контакты, объединяясь в сеть, или кластеры (кардиомиоциты и симпласты скелетной мышечной ткани). В клетке митохондрии выполняют функцию дыхания. Клеточное дыхание - это последовательность реакций, с помощью которых клетка использует энергию связей органических молекул для синтеза макроэргических соединений типа АТФ. Образующиеся внутри митохондрии молекулы АТФ переносятся наружу, обмениваясь на молекулы АДФ, находящиеся вне митохондрии. В живой клетке митохондрии могут передвигаться с помощью элементов цитоскелета. На ультрамикроскопическом уровне стенка митохондрии состоит из двух мембран - наружной и внутренней. Наружная мембрана имеет относительно ровную поверхность, внутренняя - образует направленные в центр складки, или кристы. Между наружной и внутренней мембранами возникает неширокое (около 15 нм) пространство, которое называется наружной камерой митохондрии; внутренняя мембрана ограничивает внутреннюю камеру. Содержимое наружной и внутренней камер митохондрии различно, и так же, как и сами мембраны, существенно отличается не только по рельефу поверхности, но и по ряду биохимических и функциональных признаков. Наружная мембрана по химическому составу и свойствам близка к другим внутриклеточным мембранам и плазмолемме.
Ее характеризует высокая проницаемость, благодаря наличию гидрофильных белковых каналов. Эта мембрана имеет в своем составе рецепторные комплексы, распознающие и связывающие вещества, поступающие в митохондрию. Ферментный спектр наружной мембраны небогат: это ферменты метаболизма жирных кислот, фосфолипидов, липидов и др. Главной функцией наружной мембраны митохондрии является отграничение органеллы от гиалоплазмы и транспорт необходимых для осуществления клеточного дыхания субстратов. Внутренняя мембрана митохондрий в большинстве клеток тканей различных органов формирует кристы в виде пластин (ламеллярные кристы), что значительно увеличивает площадь поверхности внутренней мембраны. В последней 20-25 % всех белковых молекул составляют ферменты дыхательной цепи и окислительного фосфорилирования. В эндокринных клетках надпочечников и половых желез митохондрии участвуют в синтезе стероидных гормонов. В этих клетках митохондрии имеют кристы в виде трубочек (тубул), упорядоченно расположенных в определенном направлении. Поэтому кристы митохондрий в стероидпродуцирующих клетках названных органов именуются тубулярными. Матрикс митохондрии, или содержимое внутренней камеры, представляет собой гелеобразную структуру, содержащую около 50 % белков. Осмиофильные тельца, описанные при электронной микроскопии, - это резервы кальция. Матрикс содержит ферменты цикла лимонной кислоты, катализирующие окисление жирных кислот, синтез рибосом, ферменты, участвующие в синтезе РНК и ДНК. Общее число ферментов превышает 40. Помимо ферментов, матрикс митохондрии содержит митохондриальную ДНК (митДНК) и митохондриальные рибосомы. Молекула митДНК имеет кольцевидную форму. Возможности внутримитохондриального белкового синтеза ограничены - здесь синтезируются транспортные белки митохондриальных мембран и некоторые ферментные белки, участвующие в фосфорилировании АДФ. Все остальные белки митохондрии кодируются ядерной ДНК, и их синтез осуществляется в гиалоплазме, и в дальнейшем они транспортируются в митохондрию. Жизненный цикл митохондрий в клетке короткий, поэтому природа наделила их двойственной системой воспроизводства - помимо деления материнской митохондрии, возможно образование нескольких дочерних органелл путем почкования.
Внешняя мембрана
Внутренняя мембрана
Матрикс
м-на, матрикс, кристы
. она имеет ровные контуры, не образует впячиваний или складок. На нее приходится около 7% от площади всех клеточных мембран. Ее толщина около 7 нм, она не бывает связана ни с какими другими мембранами цитоплазмы и замкнута сама на себя, так что представляет собой мембранный мешок. Наружнюю мембрану от внутренней отделяет межмембранное пространство
шириной около 10-20 нм. Внутренняя мембрана (толщиной около 7 нм) ограничивает собственно внутреннее содержимое митохондрии,
ее матрикс или митоплазму. Характерной чертой внутренней мембраны митохондрий является их способность образовывать многочисленные впячивания внутрь митохондрий. Такие впячивания чаще всего имеют вид плоских гребней, или крист. Расстояние между мембранами в кристе составляет около 10-20 нм. Часто кристы могут ветвиться или образовывать пальцевидные отростки, изгибаться и не иметь выраженной ориентации. У простейших, одноклеточных водорослей, в некоторых клетках высших растений и животных выросты внутренней мембраны имеют вид трубок (трубчатые кристы).
Матрикс митохондрий имеет тонкозернистое гомогенное строение, в нем иногда выявляются тонкие собранные в клубок нити (около 2-3 нм) и гранулы около 15-20нм. Теперь стало известно, что нити матрикса митохондрий представляют собой молекулы ДНК в составе митохондриального нуклеоида, а мелкие гранулы – митохондриальные рибосомы.
Функции митохондрий
1. В митохондриях происходит синтез ATP (см. Окислительное фосфорилирование)
PH межмембранного пространства ~4, pH матрикса ~8 | содержание белков в м: 67% - матрикс, 21% -наруж м-на, 6% - внутр м-на и 6% - в межм-ном пр-ве
Хандриома
– единая система митохондрий
наружная м-на: порины-поры позволяют проходить до 5 kD | внутренняя м-на: кардиолипин-делает непроницаемой м-ну для ионов |
межм-ное пр-во: группы ферментов фосфорилируют нуклеотиды и сахара нуклеотидов
внутренняя м-на:
матрикс: метаболические ферменты – окисление липидов, окисление углеводов, цикла трикарбоновых к-т, цикла Кребса
Происхождение от бактерий: амеба Pelomyxa palustris единств из эукариот не содержит м., живет в симбиозе с аэробными бактериями | собственная ДНК | схожие с бактериями оx процессы
Митохондриальная ДНК
Деление миохондрий
реплицируется
в интерфазе | репликация не связана с S-фазой | во время кл цикла митох один раз делятся надвое, образуя перетяжку, перетяжка сначала на внутр м-не | ~16,5 kb | кольцевая, кодирует 2 рРНК 22 тРНК и 13 белков |
транспорт белков: сигнальный пептид | амфифильный завиток | митохондриальный распознающий рецептор |
Окислительное фосфорилирование
Цепь переноса электронов
АТР-синтаза
в кл печени, м живут ~20 дней деление митохондрий путем образования перетяжки
16569пн=13белков,22тРНК,2pРНК | гладкая внешняя м-на (порины – проницаемость белков до 10 кДа) складчатая внутренняя (кристы) м-на (75% -белков: транспортные белки-переносчики, ф-ты, компаненты дыхат. цепи и АТФ-синтаза, кардиолипин) матрикс (обогащен ф-тами цитратного цикла) межм-ное пр-во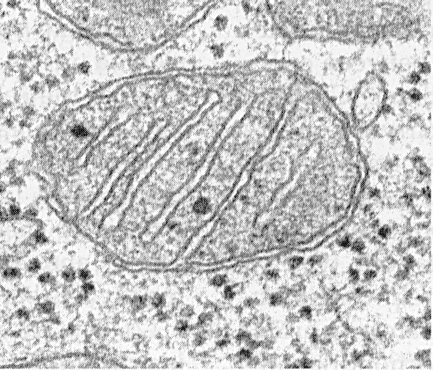
II. Митохондрии (строение и функции)
Полисомы. Синтез цитоплазматических белков
Рибосомы представляют собой мельчайшие органеллы, присутствующие в цитоплазме клетки. Несмотря на свои размеры, они являются сложными молекулярными ансамблеями, состоящими из рибосомальной РНК (р-РНК) различной длины и рибосомальных белков . В цитоплазме рибосомы встречаются в виде 2-х форм:
1. В диссоциированном состоянии (две субъединицы: малая и большая), которое свидетельствует об их неактивном статусе;
2. В ассоциированном виде – это форма их активного статуса.
Большая субъединица образуется тремя молекулами РНК, имеет форму полушара с 3 выступами, взаимодействующие с «шипиками» малой субъединицы.
Малая субъединица содержит лишь одну молекулу РНК и выглядит в виде «шапочки» с шипиками, обращёнными в сторону большой субъединицы. Ассоциация субъединиц рибосомы – это взаимодействие рельефов их поверхностей.
Функции субъединиц:
1. Малая ответственна за связывание с матричной РНК;
2. Большая – за образование полипептидной цепи.
Полисомы – это группа рибосом (от 5 до 30) связанных нитью м-РНК с образованием функционального комплекса. На нём происходит синтез цитоплазматических белков, необходимых клетке для роста, развития органелл дифференцировки.
Этапы синтеза цитоплазматических белков:
1. Выход из ядра м-РНК;
2. Сборка рибосом;
3. Образование функциональной полисомы;
4. Синтез сигнального пептида;
5. Считывание последовательности аминокислот в составе пептида сигнал-распознающей частицы (СРЧ);
6. Завершение синтеза цитоплазматического белка на полисоме. См рис. 1
Рис. 1: Схема синтеза цитоплазматических белков
II. Митохондрии (строение и функции)
Митохондрии – это система энергообеспечения клетки. На светооптическом уровне их выявляют при окраске по Альтману, они выглядят в виде зёрнышек и нитей. В цитоплазме они распределены диффузно, а в специализированных клетках сосредоточенны в участках, где имеется наибольшая потребность в энергии.
 Электронномикроскопический уровень организации митохондрии
: в ней выделяют две мембраны: наружную и внутреннюю. См. рис. 2
Электронномикроскопический уровень организации митохондрии
: в ней выделяют две мембраны: наружную и внутреннюю. См. рис. 2
Рис. 2: Схема строения митохондрии
Наружная мембрана – это мешок с относительно ровной поверхностью, она по химическому составу и свойствам близка к плазмолемме, отличается она более высокой проницаемостью и содержит ферменты метаболизма жирных кислот, фосфолипидов и липидов.
Функция:
1. Отграничение митохондрии в гиалоплазме;
2. Транспорт в митохондрию субстратов для клеточного дыхания.
Внутренняя мембрана – неровная, она формирует кристы в виде пластин (ламеллярные кристы) с увеличением площади её поверхности. Главным компонентом этой мембраны являются молекулы белков, относящиеся к ферментам дыхательной цепи, цитохромы.
На поверхности крист в некоторых клетках описывают грибовидные частицы (F 1 -частицы), в которых различают головку (9 нм) и ножку (3 нм). Считают, что именно здесь происходит синтез АТФ и АДФ.
Между наружной и внутренней мембранами образуется небольшое (около 15 – 20 нм) пространство, которое называют наружной камерой митохондрий. Внутренняя камера ограничена соответственно внутренней митохондриальной мембраной и содержит матрикс.
Матрикс митохондрий имеет гелеобразную фазу и отличается высоким содержанием белка. В нём встречаются митохондриальные гранулы – частицы диаметром 20 – 50 нм высокой электронной плотности, они содержат ионы Са 2+ и Mg 2+ . Матрикс митохондрий содержит также митохондриальные ДНК и рибосомы. На первых происходит синтез транспортных белков митохондриальных мембран и некоторых белков, участвующих в фосфолировании АДФ. ДНК здесь состоит из 37 генов и не содержит некодирующие последовательность нуклеотидов.
Функции митохондрий:
1. Обеспечение клетки энергией в виде АТФ;
2. Участие в синтезе стероидных гормонов;
3. Участие в синтезе нуклеиновых кислот;
4. Депонирование кальция.
Митохондрия – это двумембранный органоид эукариотической клетки, основная функция которого синтез АТФ – источника энергии для жизнедеятельности клетки.
Количество митохондрий в клетках не постоянно, в среднем от нескольких единиц до нескольких тысяч. Там, где процессы синтеза идут интенсивно, их больше. Также варьирует размер митохондрий и их форма (округлые, вытянутые, спиральные, чашевидные и др.). Чаще имеют округлую вытянутую форму, диаметром до 1 микрометра и длиной до 10 мкм. Могут перемещаться в клетке с током цитоплазмы или оставаться в одном положении. Перемещаются к местам, где больше всего требуется выработка энергии.
Следует иметь в виду, что в клетках АТФ синтезируется не только в митохондриях, но и в цитоплазме в процессе гликолиза . Однако эффективность этих реакций невысока. Особенность функции митохондрий в том, что в них протекают реакции не только бескислородного окисления, но и кислородный этап энергетического обмена.
Другими словами, функция митохондрий – активное участие в клеточном дыхании, к которому относят множество реакций окисления органических веществ, переноса протонов водорода и электронов, идущих с выделением энергии, которая аккумулируется в АТФ.
Ферменты митохондрий
Ферменты транслоказы внутренней мембраны митохондрий осуществляют активный транспорт АДФ и АТФ.
В структуре крист выделяют элементарные частицы, состоящие из головки, ножки и основания. На головках, состоящих из фермента АТФазы , происходит синтез АТФ. АТФаза обеспечивает сопряжение фосфорилирования АДФ с реакциями дыхательной цепи.
Компоненты дыхательной цепи находятся в основании элементарных частиц в толще мембраны.
В матриксе находится большая часть ферментов цикла Кребса и окисления жирных кислот.
В результате активности электротранспортной дыхательной цепи ионы водорода поступают в нее из матрикса, а высвобождаются на наружной стороне внутренней мембраны. Это осуществляют определенные мембранные ферменты. Разница в концентрации ионов водорода по разные стороны мембраны приводит к возникновению градиента pH.
Энергию для поддержания градиента поставляет перенос электронов по дыхательной цепи. Иначе ионы водорода диффундировали бы обратно.
Энергия градиента pH используется для синтеза АТФ из АДФ:
АДФ + Ф = АТФ + H 2 O (реакция обратима)
Образующаяся вода ферментативно удаляется. Это, наряду с другими факторами, облегчает протекание реакции слева направо.
Митохондрии или хондриосомы (от греч. mitos - нить, chondrion - зернышко, soma - тельце) представляют собой гранулярные или нитевидные органеллы, присутствующие в цитоплазме простейших, растений и животных. Митохондрии можно наблюдать в живых клетках, так как они обладают достаточно высокой плотностью. В живых клетках митохондрии могут двигаться, перемещаться, сливаться друг с другом.
У разных видов размеры митохондрий очень непостоянны, так же как изменчива их форма (рис. 199). Все же у большинства клеток толщина этих структур относительно постоянна (около 0,5 мкм), а длина колеблется, достигая у нитчатых форм до 7-60 мкм.
Изучение величины и числа митохондрий не такое простое дело. Это связано с тем, что размеры и число митохондрий, которые видны на ультратонких срезах, не соответствуют реальности.
Обычные же подсчеты показывают, что на печеночную клетку приходится около 200 митохондрий. Это составляет более 20% от общего объема цитоплазмы и около 30-35% от общего количества белка в клетке. Площадь поверхности всех митохондрий печеночной клетки в 4-5 раз больше поверхности ее плазматической мембраны. Больше всего митохондрий в ооцитах (около 300000) и у гигантской амебы Chaos chaos (до 500000).
В клетках зеленых растений число митохондрий меньше, чем в клетках животных, так как часть их функций могут выполнять хлоропласты .
Локализация митохондрии в клетках различна. Обычно митохондрии скапливаются вблизи тех участков цитоплазмы , где возникает потребность в АТФ, образующейся в митохондриях. Так, в скелетных мышцах митохондрии находятся вблизи миофибрилл. В сперматозоидах митохондрии образуют спиральный футляр вокруг оси жгутика; вероятно, это связано с необходимостью использования АТФ для движения хвоста сперматозоида. Аналогичным образом у простейших и в других клетках, снабженных ресничками, митохондрии локализуются непосредственно под клеточной мембраной у основания ресничек, для работы которых необходим АТФ. В аксонах нервных клеток митохондрии располагаются около синапсов, где происходит процесс передачи нервного импульса. В секреторных клетках, которые синтезируют большие количества белков, митохондрии тесно связаны с зонами эргастоплазмы; вероятно, они поставляют АТФ для активации аминокислот и синтеза белка на рибосомах .
Ультраструктура митохондрий.
Митохондрии независимо от их величины или формы имеют универсальное строение, их ультраструктура однообразна. Митохондрии ограничены двумя мембранами (рис. 205). Наружняя митохондриальная мембрана отделяет ее от гиалоплазмы , она имеет ровные контуры, не образует впячиваний или складок, толщина составляет около 7 нм. На нее приходится около 7% от площади всех клеточных мембран. Мембрана не связана ни с какими другими мембранами цитоплазмы, замкнута сама на себя и представляет собой мембранный мешок. Наружнюю мембрану от внутренней отделяет межмембранное пространство шириной около 10-20 нм. Внутренняя мембрана (толщиной около 7 нм) ограничивает собственно внутреннее содержимое митохондрии, ее матрикс или митоплазму. Внутренняя мембрана митохондрий образовывает многочисленные впячивания внутрь митохондрий. Такие впячивания чаще всего имеют вид плоских гребней, или крист.
Общая поверхность внутренней мембраны митохондрии в печеночной клетке составляет примерно треть поверхности всех клеточных мембран. Митохондрии клеток сердечной мышцы содержат втрое больше крист, чем печеночные митохондрии что отражает различия в функциональных нагрузках митохондрий разных клеток. Расстояние между мембранами в кристе составляет около 10-20 нм.
Митохондриальные кристы, отходящие от внутренней мембраны и простирающиеся в сторону матрикса, не перегораживают полностью полость митохондрии и не нарушают непрерывности заполняющего ее матрикса.
Ориентация крист по отношению к длинной оси митохондрии различна для разных клеток. Ориентация может быть перпендикулярная (клетки печени, почек) крист; в сердечной мышце наблюдается продольное расположение крист. Кристы могут ветвиться или образовывать пальцевидные отростки, изгибаться и не иметь выраженной ориентации (рис. 208). У простейших, одноклеточных водорослей, в некоторых клетках высших растений и животных выросты внутренней мембраны имеют вид трубок (трубчатые кристы).
Матрикс митохондрий имеет тонкозернистое гомогенное строение, в котором выявляются молекулы ДНК в виде тонких собранных в клубок нитей (около 2-3 нм) и митохондриальные рибосомы имеющие форму гранул размером около 15-20 нм. Места отложения солей магния и кальция в матриксе образуют крупные (20-40 нм) плотные гранулы.
Функции митохондрий.
Митохондрии осуществляют синтез АТФ, происходящий в результате процессов окисления органических субстратов и фосфорилирования АДФ.
Начальные этапы окисления углеводов называются анаэробным окислением, или гликолизом и происходят в гиалоплазме и не требуют участия кислорода. Субстратом окисления при анаэробном получении энергии служат гексозы и в первую очередь глюкоза; некоторые бактерии обладают свойством извлекать энергию, окисляяя пентозы, жирные кислоты или аминокислоты.
В глюкозе количество потенциальной энергии, заключенной в связях между атомами С, Н и О, составляет около 680 ккал на 1 моль (т.е. на 180 г глюкозы).
В живой клетке это огромное количество энергии освобождается в виде ступенчатого процесса, управляемого целым рядом окислительных ферментов, и не связано с переходом энергии химической связи в тепло, как при горении, а переходит в макроэнергетическую связь в молекулах АТФ, которые синтезируются при использовании освобождающейся энергии из АДФ и фосфата.
Образовавшиеся в результате гликолиза триозы, и в первую очередь пировиноградная кислота, в митохондриях вовлекаются в дальнейшее окисление. При этом происходит использование энергии расщепления всех химических связей, что приводит к выделению СО 2, к потреблению кислорода и синтезу большого количества АТФ. Эти процессы связаны с окислительным циклом трикарбоновых кислот и с дыхательной цепью переноса электронов, где происходит фосфорилирование АДФ и синтез клеточного “топлива”, молекул АТФ (рис. 209).
В цикле трикарбоновых кислот (цикл Кребса , или цикл лимонной кислоты) образовавшийся в результате гликолиза пируват сначала теряет молекулу СО 2 и, окисляясь до ацетата (двууглеродное соединение), соединяется с коферментом А. Затем ацетилкоэнзим А, соединяясь с оксалацетатом (четырехуглеродное соединение), образует шестиуглеродный цитрат (лимоную кислоту). Затем происходит цикл окисления этого шестиуглеродного соединения до четырехуглеродного оксалацетата, снова связывание с ацетилкоэнзимом А, и затем цикл повторяется. При этом окислении выделяются две молекулы СО 2, а электроны, освободившиеся при окислении, переносятся на акцепторные молекулы коферментов (NAD-никотинамидадениндинуклеотид), которые вовлекают их далее в цепь переноса электронов. Следовательно, в цикле трикарбоновых кислот нет самого синтеза АТФ, а идет окисление молекул, перенос электронов на акцепторы и выделение СО 2 . Все описанные выше события внутри митохондрий происходят в их матриксе.
Окисление исходного субстрата приводит к выделению СО 2 и воды, но при этом не выделяется тепловая энергия, как при горении, а образовываются молекулы АТФ. Они синтезируются другой группой белков, не связанных прямо с окислением. Во внутренних митохондриальных мембранах на поверхности мембран, смотрящих в матрикс, располагаются крупные белковые комплексы, ферменты, АТФ-синтетазы. В электронном микроскопе они видны в виде так называемых “грибовидных” телец сплошь выстилающие поверхность мембран, смотрящую в матрикс. Тельца имеют как бы ножку и головку, диаметром 8-9 нм. Следовательно, во внутренних мембранах митохондрий локализованы ферменты как окислительной цепи, так и ферменты синтеза АТФ (рис. 201б).
Дыхательная цепь - это главная система превращения энергии в митохондриях. Здесь происходит последовательное окисление и восстановление элементов дыхательной цепи, в результате чего высвобождается небольшими порциями энергия. За счет этой энергии в трех точках цепи из АДФ и фосфата образуется АТФ. Поэтому говорят, что окисление (перенос электронов) сопряжено с фосфорилированием (АДФ + Фн →АТФ, т.е. происходит процесс окислительного фосфорилирования.
Выделяющаяся при транспорте электронов энергия запасается в виде градиента протонов на мембране. Оказалось, что при переносе электронов в митохондриальной мембране каждый комплекс дыхательной цепи направляет свободную энергию окисление на перемещение протонов (положительных зарядов) через мембрану, из матрикса в межмембранное пространство, что приводит к образованию разности потенциалов на мембране: положительные заряды преобладают в межмембранном пространстве, а отрицательные - со стороны матрикса митохондрий. При достижении разности потенциалов (220 мВ) белковый комплекс АТФ-синтетазы начинает транспортировать протоны обратно в матрикс, при этом превращает одну форму энергии в другую: образует АТФ из АДФ и неорганического фосфата. Так происходит сопряжение окислительных процессов с синтетическим, с фосфорилированием АДФ. Пока происходит окисление субстратов, пока происходит перекачка протонов через внутреннюю митохондриальную мембрану - идет сопряженный с этим синтез АТФ, т.е. происходит окислительное фосфорилирование.
Эти два процесса можно разобщить. При этом продолжается перенос электронов, как и окисление субстрата, но синтеза АТФ не происходит. В этом случае энергия, освобождающаяся при окислении переходит в тепловую энергию.
Окислительное фосфорилирование у бактерий .
У прокариотических клеток, способных к окислительному фосфорилированию, элементы цикла трикарбоновых кислот локализованы прямо в цитоплазме, а ферменты дыхательной цепи и фосфорилирования связаны с клеточной мембраной, с ее выпячиваниями, выступающими внутрь цитоплазмы, с так называемыми мезосомами (рис. 212). Надо отметить, что такие бактериальные мезосомы могут быть связаны не только с процессами аэробного дыхания, но и у некоторых видов участвовать в делении клеток, в процессе распределения ДНК по новым клеткам, в образовании клеточной стенки и т.д.
На плазматической мембране в мезосомах некоторых бактерий осуществляются сопряженные процессы как окисления так и синтеза АТФ. В электронном микроскопе во фракциях плазматических мембран бактерий обнаружены сферические частицы, аналогичные тем, которые были найдены в митохондриях эукариотических клеток. Таким образом, у бактериальных клеток, способных к окислительному фосфорилированию, плазматическая мембрана выполняет роль, аналогичную внутренней мембране митохондрий эукариотических клеток.
Увеличение числа митохондрий.
Митохондрии могут увеличивать свою численность особенно при делении клеток или при увеличении функциональной нагрузки клетки. Происходит постоянное обновление митохондрий. Например, в печени средняя продолжительность жизни митохондрий составляет около 10 дней.
Увеличение числа митохондрий происходит путем роста и деления предшествующих митохондрий. Это предположение было впервые высказано Альтманом (1893), описавшим митохондрии под термином “биобласты”. Удается наблюдать прижизненно деление, фрагментацию длинных митохондрий на более короткие путем перетяжки, что напоминает бинарный способ деления бактерий.
Реальное увеличение числа митохондрий путем деления установлено при изучении поведения митохондрий в живых клетках культуры ткани. В течение клеточного цикла митохондрии вырастают до нескольких мкм, а затем фрагментируются, делятся на более мелкие тельца.
Митохондрии могут сливаться друг с другом и размножаться по принципу: митохондрия от митохондрии.
Авторепродукция митохондрий.
Двумембранные органеллы обладают полной системой авторепродукции. В митохондриях и пластидах существует ДНК, на которой синтезируются информационные, трансферные и рибосомные РНК и рибосомы, осуществляющие синтез митохондриальных и пластидных белков. Однако, эти системы, хотя и автономны, но ограничены по своим возможностям.
ДНК в митохондриях представляет собой циклические молекулы без гистонов и тем самым напоминают бактериальные хромосомы. Размер их составляет около 7 мкм, в одну циклическую молекулу митохондрий животных входит 16-19 тыс. нуклеотидных пар ДНК. У человека митохондриальная ДНК содержит 16,5 тыс. н.п., она полностью расшифрована. Найдено, что митохондральная ДНК различных объектов очень однородна, отличие их заключается лишь в величине интронов и нетранскрибируемых участков. Все митохондриальные ДНК представляют множественные копии, собранными в группы, кластеры. Так в одной митохондрии печени крысы может содержаться от 1 до 50 циклических молекул ДНК. Общее же количество митохондриальной ДНК на клетку составляет около одного процента. Синтез митохондриальных ДНК не связан с синтезом ДНК в ядре.
Так же как и у бактерий митохондральная ДНК собрана в отдельную зону - нуклеоид, его размер составляет около 0, 4 мкм в диаметре. В длинных митохондриях может быть от 1 до 10 нуклеоидов. При делении длинной митохондрии от нее отделяется участок, содержащий нуклеоид (сходство с бинарным делением бактерий). Количество ДНК в отдельных нуклеоидах митохондрий может колебаться в 10 раз в зависимости от типа клеток.
В некоторых культурах в клетках от 6 до 60% митохондрий не имеют нуклеоида, что может объясняться тем, что деление этих органелл скорее связано с фрагментацией, а не с распределением нуклеоидов.
Как уже говорилось, митохондрии могут как делиться, так и сливаться друг с другом. При слиянии митохондрий друг с другом может происходить обмен их внутренними компонентами.
Важно подчеркнуть, что рРНК и рибосомы митохондрий и цитоплазмы резко отличны. Если в цитоплазме обнаруживаются 80s рибосомы, то рибосомы митохондрий растительных клеток принадлежат к 70s рибосомам (состоят из 30s и 50s субъединиц, содержат 16s и 23s РНК, характерные для прокариотических клеток), а в митохондриях клеток животных обнаружены более мелкие рибосомы (около 50s).
Рибосомные РНК митохондрий синтезируются на митохондриальных ДНК. В митоплазме на рибосомах идет синтез белков. Он прекращается, в отличие от синтеза на цитоплазматических рибосомах, при действии антибиотика хлорамфеникола, подавляющего синтез белка у бактерий.
На митохондриальном геноме синтезируются 22 транспортные РНК. Триплетный код митохондриальной синтетической системы отличен от такового, используемого в гиалоплазме. Несмотря на наличие казалось бы всех компонентов, необходимых для синтеза белков, небольшие молекулы митохондриальной ДНК не могут кодировать все митохондриальные белки, только лишь их небольшую часть. Так ДНК размером 15 тыс.н.п. может кодировать белки с суммарным молекулярным весом около 6х10 5 . В это же время суммарный молекулярный вес белков частицы полного дыхательного ансамбля митохондрии достигает величины около 2х10 6 .
Если учесть, что кроме белков окислительного фосфорилирования в митохондрии входят ферменты цикла трикарбоновых кислот, ферменты синтеза ДНК и РНК, ферменты активации аминокислот и другие белки, то видно, что, для того чтобы кодировать эти многочисленные белки и рРНК и тРНК, количества генетической информации в короткой молекуле митохондриальной ДНК явно не хватает. Расшифровка нуклеотидной последовательности митохондриальной ДНК человека показала, что она кодирует всего лишь 2 рибосомные РНК, 22 трансферных РНК и всего 13 различных полипептидных цепей.
В настоящее время доказано, что большая часть белков митохондрий находится под генетическим контролем со стороны клеточного ядра и синтезируется вне митохондрий. Большинство митохондриальных белков синтезируется на рибосомах в цитозоле. Эти белки имеют специальные сигнальные последовательности, которые узнаются рецепторами на внешней мембране митохондрий. Эти белки могут встраиваться в них (см. аналогию с мембраной пероксисом), а затем перемещаться на внутреннюю мембрану. Этот перенос происходит в точках контакта наружной и внутренней мембран, где такой транспорт отмечен. Большинство липидов митохондрий так же синтезируются в цитоплазме.
Все это говорит о эндосимбиотическом происхождении митохондрий, о том, что митохондрии представляют собой организмы типа бактерий, находящиеся в симбиозе с эукариотический клеткой.
Хондриом.
Совокупность всех митохондрий в одной клетке называется хондриомом. Она может быть различной в зависимости от типа клеток. Во многих клетках хондриом состоит из разрозненных многочисленных митохондрий, равномерно расположенных по всей цитоплазме или локализуются группами в местах интенсивной траты АТФ. В обоих этих случаях митохондрии функционируют поодиночке, их кооперативная работа, возможно, координируется какими-то сигналами из цитоплазмы. Существует и совершенно иной тип хондриома, когда вместо мелких одиночных разрозненных митохондрий в клетке располагается одна гигантская разветвленная митохондрия.
Такие митохондрии встречаются у одноклеточных зеленых водорослей (например у Chlorella). Они образуют сложную митохондриальную сеть или митохондриальный ретикулум (Reticulum miyochondriale). Согласно хемоосмотической теории биологический смысл появления такой гигантской разветвленной митохондриальной структуры, объединенной в одно целое своими внешними и внутренними мембранами заключается в том, что в любой точке поверхности внутренней мембраны такой разветвленной митохондрии может идти синтез АТФ, который будет поступать в любую точку цитоплазмы, где в этом есть необходимость.
В случае гигантских разветвленных митохондрий в любой ее точке может на внутренней мембране накопиться потенциал, достаточный для того, чтобы начался синтез АТФ. С этих позиций митохондриальный ретикулум представляет собой как бы электрический проводник, кабель, соединяющий отдаленные точки такой системы. Митохондриальный ретикулум оказался очень полезным не только для мелких подвижных клеток, таких как хлорелла, но и для более крупных структурных единиц таких как, например, миофибриллы в скелетных мышцах.
Известно, что скелетные мышцы состоят из массы мышечных волокон, симпластов, содержащих множество ядер. Длина таких мышечных волокон достигает 40 мкм, при толщине 0,1 мкм - это гигантская структура, содержащая великое множество миофибрилл, все из которых сокращаются одновременно, синхронно. Для сокращения к каждой единице сокращения, к миофибрилле, доставляется большое количество АТФ, которых обеспечивают митохондрии на уровне z-дисков. На продольных ультратонких срезах скелетных мышц в электронном микроскопе видны многочисленные округлые мелкие сечения митохондрий, располагающихся в соседстве с саркомерами. Мышечные митохондрии представляют собой не мелкие шарики или палочки, а как бы паукообразные структуры, отростки которых ветвятся и простираются на большие расстояния, иногда через весь поперечник мышечного волокна.
При этом разветвления митохондрий окружают каждую миофибриллу в мышечном волокне, снабжая их АТФ, необходимого для мышечного сокращения. Следовательно, в плоскости z-диска митохондрии представляют типичный митохондриальный ретикулум. Такой пласт или этаж митохондриального ретикулума повторяется дважды на каждый саркомер, а все мышечное волокно имеет тысячи поперечно расположенных “поэтажных” пластов митохондриального ретикулума. Между “этажами” вдоль миофибрилл располагаются нитчатые митохондрии, соединяющие эти митохондриальные пласты. Таким образом создана трехмерная картина митохондриального ретикулума, проходящего через весь объем мышечного волокна.
Далее было установлено, что между ответвлениями митохондриального ретикулума и нитевидными продольными митохондриями существуют специальные межмитохондриальные соединения или контакты (ММК). Они образованы плотно прилегающими наружными митохондриальными мембранами контактирующих митохондрий, межмембранное пространство и мембраны в этой зоне имеют повышенную электронную плотность. Через эти специальные образования происходит функциональное объединение соседних митохондрий и митохондриальных ретикулумов в единую, кооперативную энергетическую систему. Все миофибриллы в мышечном волокне сокращаются синхронно по всей их длине, следовательно, и поступление АТФ на любом участке этой сложной машины тоже должно происходить синхронно, а это может происходить лишь в том случае, если огромное количество разветвленных митохондрий-проводников будет связано друг с другом с помощью контактов.
О том, что межмитоходриальные контакты (ММК) участвуют в энергетическом объединении митохондрий друг с другом удалось на кардиомиоцитах, клетках сердечных мышц.
Хондриом клеток сердечной мышцы не образует ветвящихся структур, а представлен множеством небольших вытянутых митохондрий, располагающихся без особого порядка между миофибриллами. Однако, все соседние митохондрии стыкуются друг с другом с помощью митохондриальных контактов такого же типа, как в скелетной мышце, только их число очень велико: в среднем на одну митохондрию приходится 2-3 ММК, которые связывают митохондрии в единую цепь, где каждым звеном такой цепи (Streptio mitochondriale) является отдельная митохондрия.
Оказалось, что межмитохондриальные контакты (ММК), как обязательная структура сердечных клеток обнаружены в кардиомиоцитах как желудочков, так и предсердий всех позвоночных животных: млекопитающих, птиц, пресмыкающихся, амфибий и костистых рыб. Более того ММК были обнаружены (но в меньшем числе) в клетках сердца некоторых насекомых и моллюсков.
Количество ММК в кардиомиоцитах изменяется в зависимости от функциональной нагрузки на сердце. Число ММК увеличивается при повышении физических нагрузок животных и, наоборот, при падении нагрузки на сердечную мышцу происходит резкое сокращение числа ММК.

